Papers
You’ll find a complete list of papers on my CV (see About for link) and ResearchGate
-
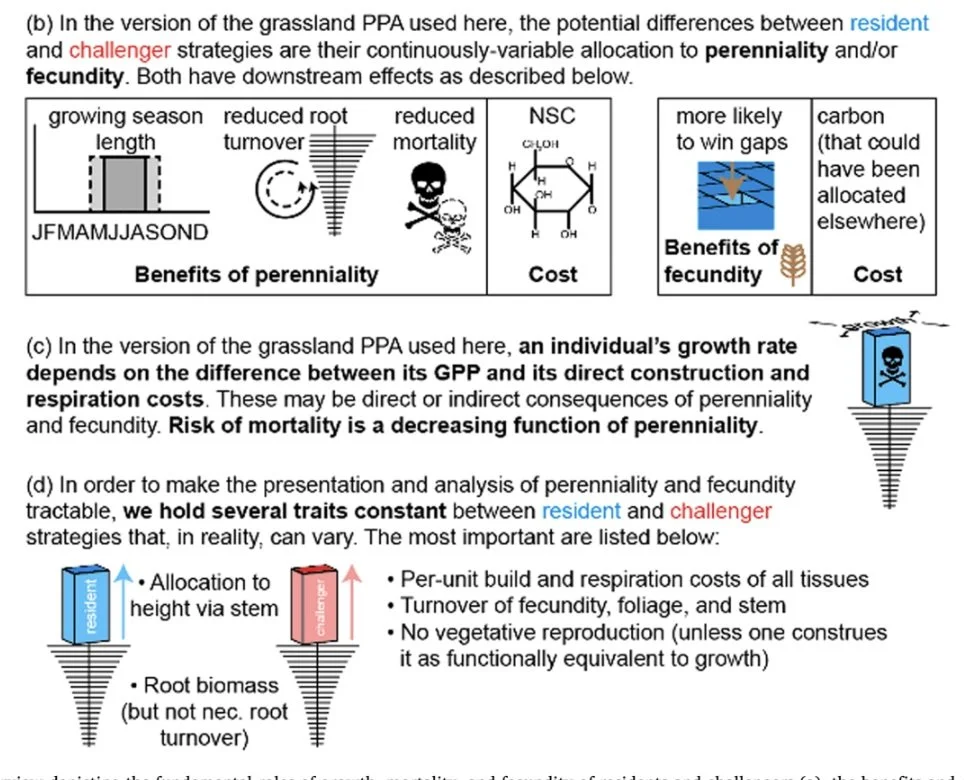
Modeling carbon allocation strategies for high-yielding perennial crops. Kilbane et al. 2025 EuroJAg
We constructed a generalizable grassland model of within-plant carbon allocation strategies with the objective of furthering understanding of the evolutionary ecology of perenniality and illuminating possibilities for breeding high-yielding, long-lived crops in the service of regenerative agriculture. The grassland Perfect Plasticity Approximation model handles space-filling, recruitment, and carbon balance to determine the within-plant allocation of carbon given trade-offs involving perenniality. We used our model to conduct (1) a game-theoretic analysis of evolutionarily stable strategies to determine the effects of natural selection on perenniality and annual reproductive yield, and (2) analyses of physiological feasibility and sensitivity to determine the range of allocational strategies that might be achievable through breeding. Model results suggest that natural selection already maximizes annual reproductive yield in plants that have adapted to the evolutionarily stable strategy in the context of constraints of carbon allocation to foliage, roots, stems, and growth, leaving little room for yield improvement without also breeding a reduction in carbon allocation elsewhere. Breeding reductions in stem mass and root mass show promise for increasing annual reproductive yield. Further research is needed, however, to understand how reductions in root or stem mass would affect a perennial’s provisioning of ecosystem services.
-
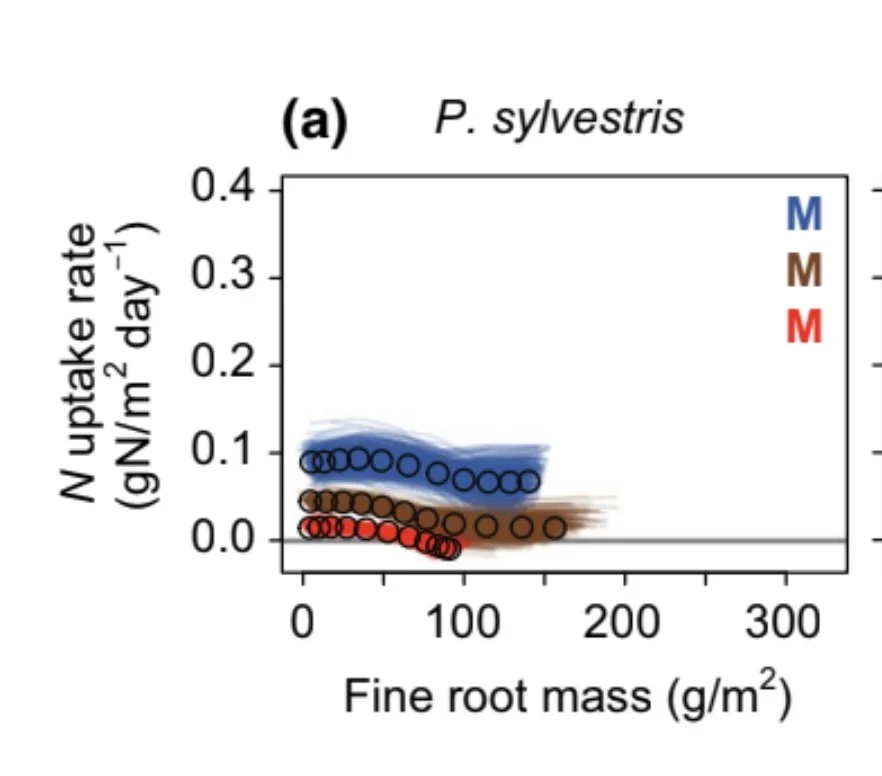
How are nitrogen availability, fine‐root mass, and nitrogen uptake related empirically? Implications for models and theory. Dybzinski et al. 2019 GCB
Understanding the effects of global change in terrestrial communities requires an understanding of how limiting resources interact with plant traits to affect productivity. Here, we focus on nitrogen and ask whether plant community nitrogen uptake rate is determined (i) by nitrogen availability alone or (ii) by the product of nitrogen availability and fine-root mass. Surprisingly, this is not empirically resolved. We performed controlled microcosm experiments and reanalyzed published pot experiments and field data to determine the relationship between community-level nitrogen uptake rate, nitrogen availability, and fine-root mass for 46 unique combinations of species, nitrogen levels, and growing conditions. We found that plant community nitrogen uptake rate was unaffected by fine-root mass in 63% of cases and saturated with fine-root mass in 29% of cases (92% in total). In contrast, plant community nitrogen uptake rate was clearly affected by nitrogen availability. The results support the idea that although plants may over-proliferate fine roots for individual-level competition, it comes without an increase in community-level nitrogen uptake. The results have implications for the mechanisms included in coupled carbon-nitrogen terrestrial biosphere models (CN-TBMs) and are consistent with CN-TBMs that operate above the individual scale and omit fine-root mass in equations of nitrogen uptake rate but inconsistent with the majority of CN-TBMs, which operate above the individual scale and include fine-root mass in equations of nitrogen uptake rate. For the much smaller number of CN-TBMs that explicitly model individual-based belowground competition for nitrogen, the results suggest that the relative (not absolute) fine-root mass of competing individuals should be included in the equations that determine individual-level nitrogen uptake rates. By providing empirical data to support the assumptions used in CN-TBMs, we put their global climate change predictions on firmer ground.
-
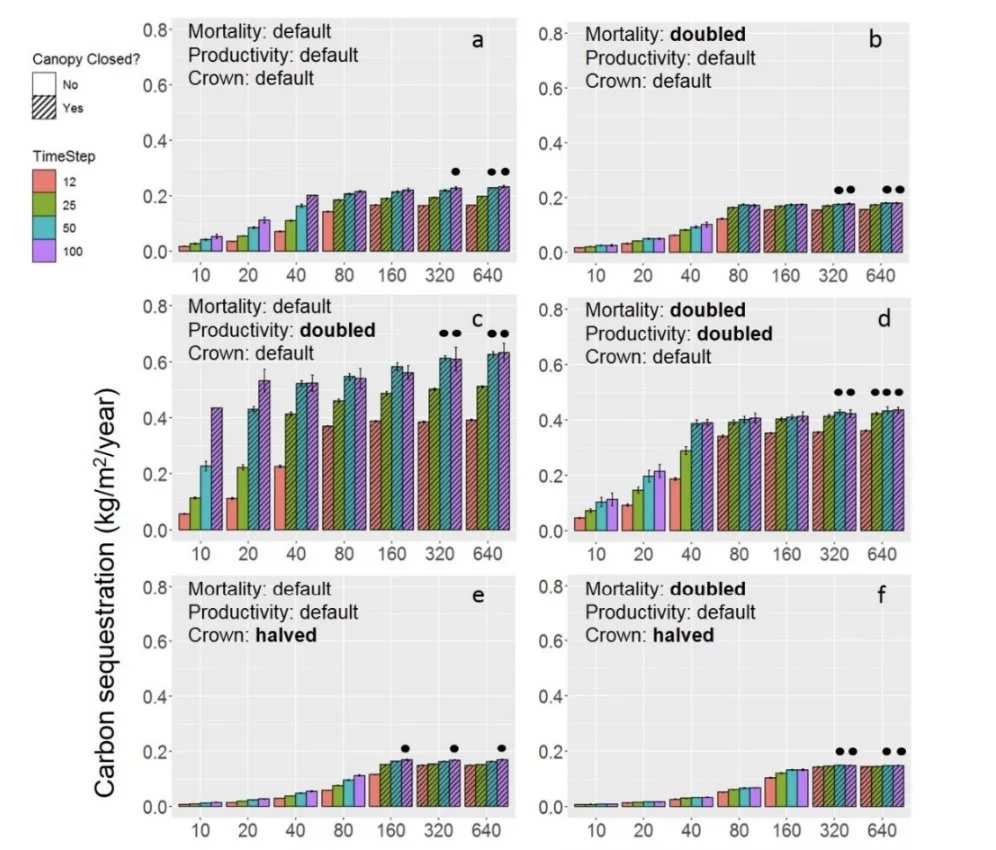
Modeling Management-Relevant Urban Forest Stand Characteristics to Optimize Carbon Storage and Sequestration. Drolen et al. 2023 Forests
Urban forests are an essential part of adaptation and mitigation solutions for climate change. To understand the relationship between carbon storage, sequestration, and stand density in the most heavily-managed aspect of the urban forest—street trees—we modified the parameters and algorithms of a rural forest dynamics model, the Perfect Plasticity Approximation (PPA), to reflect urban street tree conditions. The main changes in the new street tree PPA are the maintenance of a prescribed stand density via management of recruitment, the possibility of crown-roof overlap, and increased mortality rates. Using the street tree PPA, we explored overall productivity, crown allometry relative to stem diameter, and mortality rate to test each mechanism’s impact on urban street tree carbon storage and sequestration across a gradient of prescribed stand density, with the goal of finding conditions in which street tree carbon storage and sequestration are optimized. We compared the qualitative trends in storage from the street tree PPA to those found in the U.S. Forest Service’s Urban Forest Inventory Analysis data. We found that carbon storage and sequestration increase with prescribed density up to a point where carbon storage and sequestration saturate. Optimized carbon storage and sequestration result from a stand with high productivity, maximized crown allometry relative to stem diameter, and a low mortality rate. These insights can be used to inform urban street tree maintenance strategies that effectively increase carbon storage and sequestration within a given city, such as focusing afforestation campaigns on adequate areas with the lowest street tree densities.
-
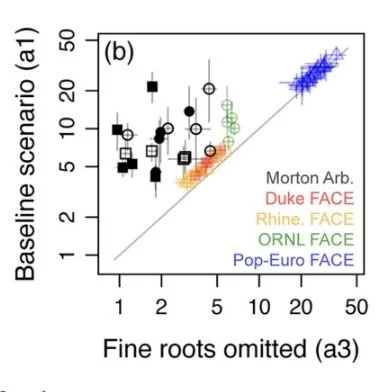
Calculating nitrogen uptake rates in forests: which components can be omitted, simplified, or taken from trait databases and which must be measured in situ? Dybzinski et al. 2024 Ecosystems
Quantifying nitrogen uptake rates across different forest types is critical for a range of ecological questions, including the parameterization of global climate change models. However, few measurements of forest nitrogen uptake rates are available due to the intensive labor required to collect in situ data. Here, we seek to optimize data collection efforts by identifying measurements that must be made in situ and those that can be omitted or approximated from databases. We estimated nitrogen uptake rates in 18 mature monodominant forest stands comprising 13 species of diverse taxonomy at The Morton Arboretum in Lisle, IL USA. We measured all nitrogen concentrations, foliage allocation, and fine root biomass in situ. We estimated wood biomass increments by in situ stem diameter and stem core measurements combined with allometric equations. We estimated fine root turnover rates from database values. We analyzed similar published data from monodominant forest FACE sites. At least in monodominant forests, accurate estimates of forest nitrogen uptake rates appear to require in situ measurements of fine root productivity and are appreciably better paired with in situ measurements of foliage productivity. Generally, wood productivity and tissue nitrogen concentrations may be taken from trait databases at higher taxonomic levels. Careful sorting of foliage or fine roots to species is time consuming but has little effect on estimates of nitrogen uptake rate. By directing research efforts to critical in situ measurements only, future studies can maximize research effort to identify the drivers of varied nitrogen uptake patterns across gradients.
-
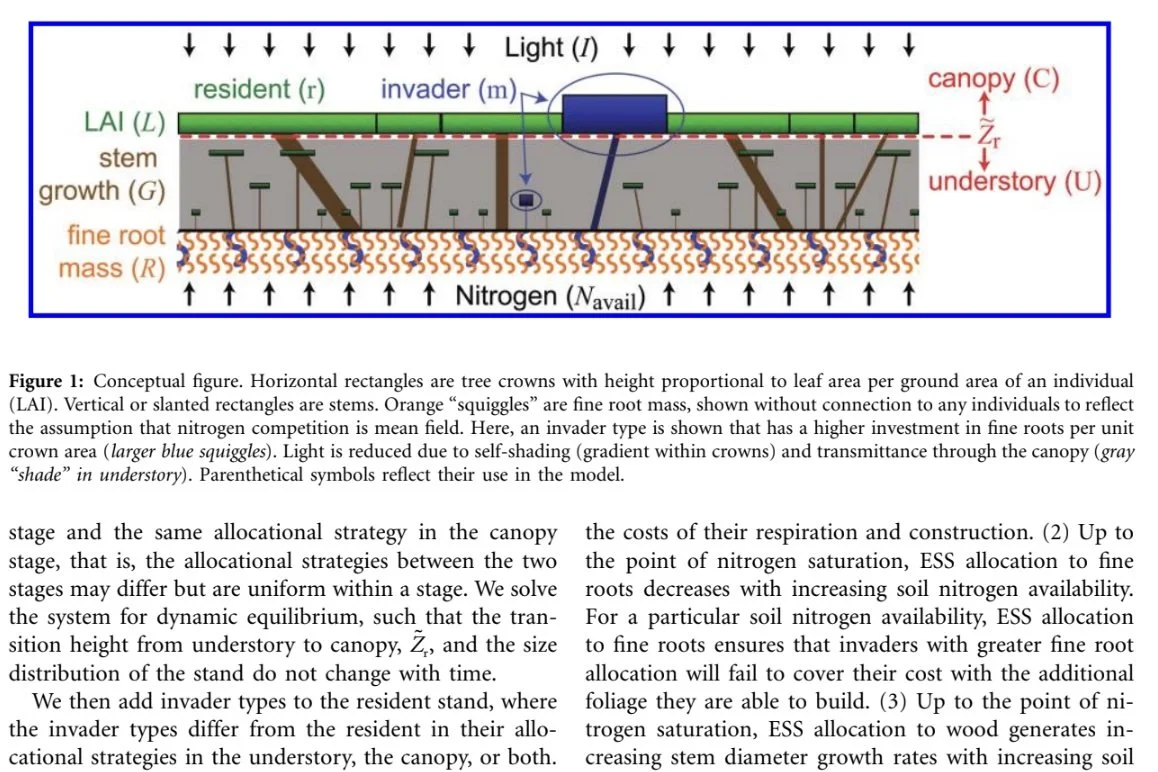
Evolutionarily Stable Strategy Carbon Allocation to Foliage, Wood, and Fine Roots in Trees Competing for Light and Nitrogen: An Analytically Tractable, Individual-Based Model and Quantitative Comparisons to Data. Dybzinski et al. 2011 AmNat
We present a model that scales from the physiological and structural traits of individual trees competing for light and nitrogen across a gradient of soil nitrogen to their community-level consequences. The model predicts the most competitive (i.e., the evolutionarily stable strategy [ESS]) allocations to foliage, wood, and fine roots for canopy and understory stages of trees growing in oldgrowth forests. The ESS allocations, revealed as analytical functions of commonly measured physiological parameters, depend not on simple root-shoot relations but rather on diminishing returns of carbon investment that ensure any alternate strategy will underperform an ESS in monoculture because of the competitive environment that the ESS creates. As such, ESS allocations do not maximize nitrogen-limited growth rates in monoculture, highlighting the underappreciated idea that the most competitive strategy is not necessarily the "best," but rather that which creates conditions in which all others are "worse. " Data from 152 stands support the model's surprising prediction that the dominant structural trade-off is between fine roots and wood, not foliage, suggesting the "root-shoot" trade-off is more precisely a "root-stem" trade-off for long-lived trees. Assuming other resources are abundant, the model predicts that forests are limited by both nitrogen and light, or nearly so.
-
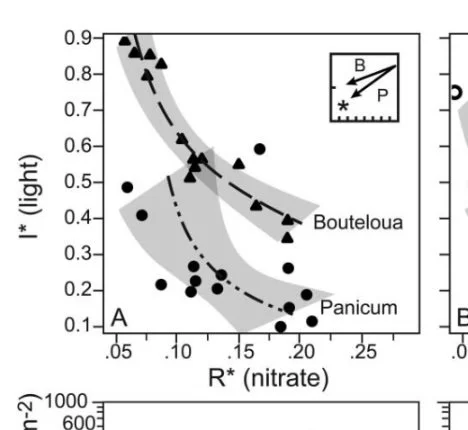
Resource use patterns predict long-term outcomes of plant competition for nutrients and light. Dybzinski & Tilman 2007 AmNat
An 11-year competition experiment among combinations of six prairie perennial plant species showed that resource competition theory generally predicted the long-term outcome of competition. We grew each species in replicated monocultures to determine its requirements for soil nitrate (R*) and light (I*). In six pairwise combinations, the species with the lower R* and I* excluded its competitor, as predicted by theory. In the remaining two pairwise combinations, one species had a lower R* and the second had a lower I*; these species pairs coexisted, although it is unclear whether resource competition alone was responsible for their coexistence. Smaller differences in R* or I* between competing species led to slower rates of competitive exclusion, and the influence of R* differences on the rate of competitive exclusion was more pronounced on low nitrogen soils while the influence of I* differences was more pronounced on high nitrogen (low light) soils. These results were not explained by differences in initial species abundances or neutrality. However, only a few of our paired species coexisted under our experimentally-imposed conditions (homogeneous soils, high seeding densities, minimal disturbance, regular water, and low herbivory levels), suggesting that other coexistence mechanisms help generate the diversity observed in natural communities.